Worn teeth are a bane to paleobiologists interested in the diets of human ancestors and other fossil primates. Although worn teeth dominate fossil assemblages, their shapes are usually not used to reconstruct the diets of extinct species. The problem is that traditional studies of primate dental functional anatomy have focused on unworn morphology. This has limited most functional analyses to only a few well-represented fossil species. This paper introduces a method to characterize and compare worn occlusal morphology in primates using laser scanning and geographic information systems technologies. A study of variably worn chimpanzee and gorilla molars indicates that differences between these species in tooth shape remain consistent at given stages of wear. Although cusp slope decreases with wear in both taxa, angularity values remain unchanged. These results indicate that African ape teeth wear in a manner that keeps them mechanically efficient for fracturing specific foods. Studies of changes in tooth shape with wear add a new dimension to dental functional anatomy, and offer a more complete picture of dental dietary adaptations. Also, given how rare unworn teeth are in the fossil record, the ability to include worn specimens in analyses opens the door to reconstructing the diets of many more extinct primate groups, allowing us to better understand the adaptive radiation of our order.
Feeding adaptations are of great interest to primatologists, whether they study living or fossil species. Diet underlies many of the behavioral and ecological differences that separate species, and so is important in defining niche, with all its implications for the ecology and evolution of extinct forms. Most attempts to reconstruct diet in fossil taxa have focused on teeth, which tend to dominate fossil assemblages and, as durable parts of the digestive system, offer a fairly direct glimpse at what past primates ate.
Researchers have recognized for the better part of a century that primate tooth shape has evolved to improve mechanical efficiency for chewing (1). Because foods differ in their fracture properties, tooth morphology differs among species adapted to different diets (2). Therefore, an understanding of relationships between tooth form and food properties has led many paleontologists to reconstruct the diets of fossil primate species by using tooth shape.
Most such studies have focused on molar shearing crest lengths (3). The summed lengths of mesiodistal crests are regressed overocclusal surface length for closely related extant species with a given diet. The position of a fossil species relative to the regression line indicates whether the lengths of its shearing crests are within the range expected for that diet. Studies of relative shearing crest length have shown that this measure readily distinguishes primates by broad diet category. Folivores, for example, have relatively longer shearing crests than frugivores, and, among frugivores, soft-fruit eaters have longer crests than hard-object specialists (4). Although such studies provide a useful baseline for the inference of aspects of diet in fossil taxa, they are limited by their reliance on unworn teeth. Given that these crests are measured as distances between cusp tips and the notches between cusps, crest lengths change with attrition, and quickly become impossible to measure as cusp tips are obliterated by wear.
Restricting studies of dental functional anatomy to unworn teeth is, however, not ideal. First, most fossil primate teeth are worn, and samples of unworn fossil teeth are often insufficient to allow statistical comparisons with extant baseline series. There are, for example, fewer than 10 unworn lower second molars in the entire collection of published australopith teeth from South Africa (5). Second, teeth wear, and natural selection does not stop when the first facets appear on an occlusal surface. We cannot hope to understand tooth form without considering that selection should favor shapes that wear in a manner designed to keep them mechanically efficient for fracturing foods. Not only should foods with different material properties select for teeth with different unworn occlusal morphologies, but also these foods should select for teeth that maintain these differences with wear, or even change their shapes in different ways as they wear (6, 7).
The need to examine and compare occlusal form in variably worn teeth calls for a new approach to dental morphometrics. This paper describes a method for characterizing and comparing tooth form in variably worn teeth by using a laser scanner to generate 3D points from the surface of a molar tooth, and geographic information systems (GIS) software to model and measure functional aspects of occlusal morphology.
GIS is a system for assembling, storing and analyzing geographically referenced information for a broad range of applications. Many GIS tools have been created to examine and model the physical surface of the Earth, and these tools offer equal potential for characterizing occlusal surfaces of primate teeth in three dimensions. Preliminary studies have already shown that mammalian tooth surfaces can be digitized by using a variety of instruments and modeled for analysis using GIS techniques (8 –11).
The study described here expands the GIS approach to dental topographic analysis to include the characterization and comparison of worn teeth. This technique is used to document functional differences in occlusal morphology between species and to determine how tooth form changes with wear. A study of chimpanzee and gorilla molars shows that cusp slopes decrease as they wear in both of these species, and that differences between taxa are maintained through the wear sequence. Cusp angularity also differs between species, but angularity does not change significantly with wear in either taxon. This finding suggests that chimpanzee and gorilla molar shapes are selected to maintain functional differences in morphology despite wear. Indeed, some aspects of occlusal form remain unchanged even as tooth cusps are worn away.
Methods
This study examined lower second molar teeth of wild-caught Gorilla gorilla gorilla(n=48) and Pan troglodytes troglodytes(n=56) specimens from the Cleveland Museum of Natural History. These teeth were chosen following previous studies of molar functional morphology (4). These taxa were chosen for two reasons. First, African apes serve as good models for under-standing dental functional morphology in early hominins and other fossil higher primates. Second, they have been reported to differ in their diets in ways likely to affect their dental morphol-ogies: gorillas are more folivorous, whereas chimpanzees are more frugivorous. The degree of dietary difference between the taxa was controlled by selection of the western lowland gorilla, which more often consumes fruits than the better-known moun-tain gorilla subspecies and is found sympatric with P. t. troglodytes(12, 13).
Only undamaged specimens that preserved sufficient morphology to identify boundaries of individual cusps were included in this study. High-resolution replicas were prepared by using conventional dental microwear casting procedures (14). Molds were made by using President’s Jet (Coltène, Mahwah, NJ) polyvinylsiloxane impression material. Casts were poured by using Epotek 301 (Epoxy Technologies, Billerica, MA), and coated with a thin layer of Magnif lux SKD-S2 Developer (Illinois Tool Works, Glenview, IL) to mitigate specimen translucency. The end result was a series of tooth replicas, reproducing surface features accurate to a fraction of a micrometer.
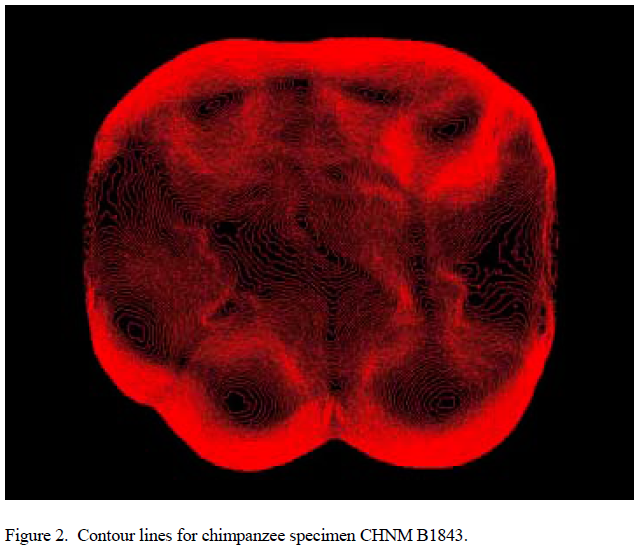
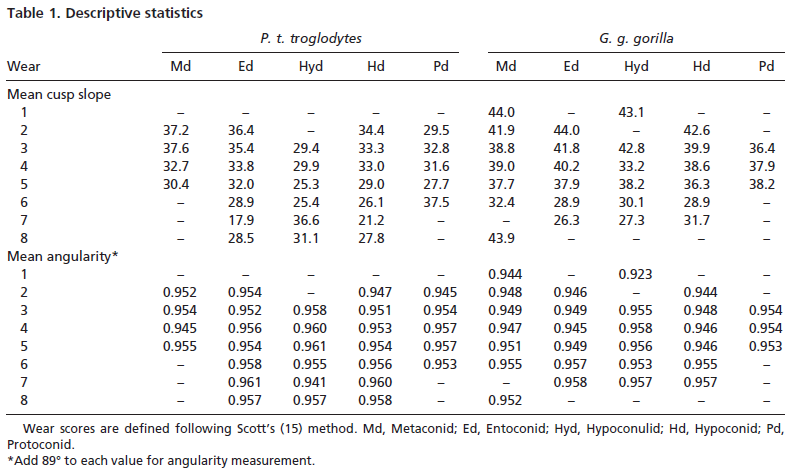
Occlusal surfaces of these replicas were scanned with amodified Surveyor 500 scanner using an RPS-150 laser (Laser Design, Minneapolis). This instrument produces a series of z values at fixed intervals in x and y along the surface of a tooth. The resolution used for this study was 25.4 um in x,y, and z. Resulting coordinate data were analyzed in ARCVIEW 3.1 with the Spatial Analyst extension (ESRI, Redlands, CA). Occlusal surfaces were interpolated by inverse distance weighting. To assure comparability of specimens and limit analysis to the functional occlusal table, each surface was cropped to the horizontal plane intersecting the lowest point in the central basin of each speci- men. Individual cusps were then delineated with the help of contour lines, and scored for wear by using Scott’s (15) method.
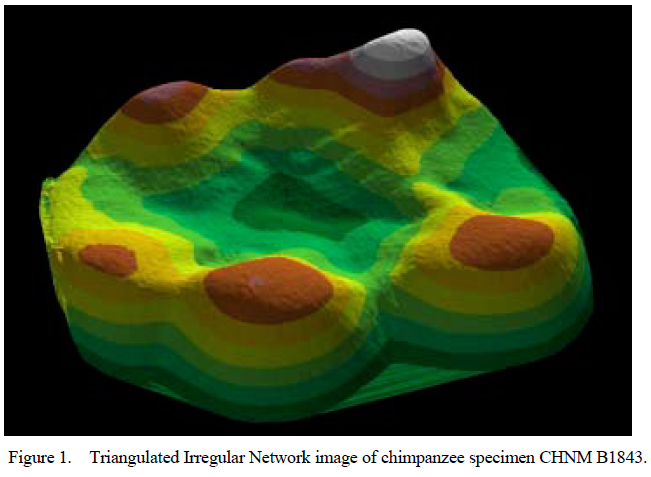
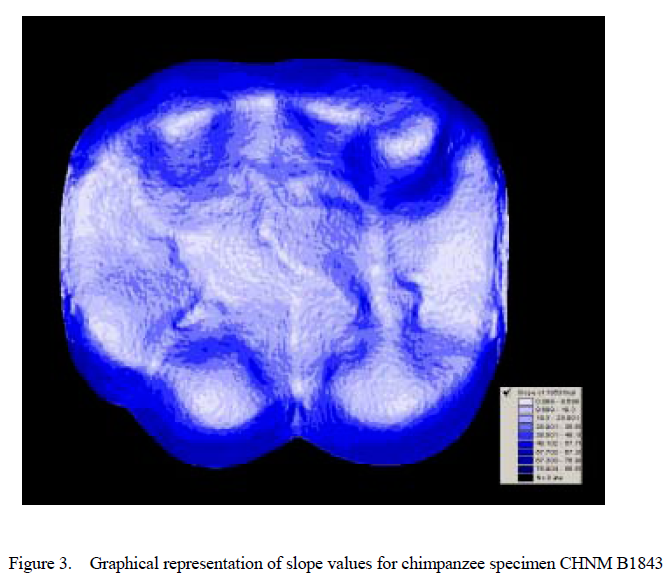
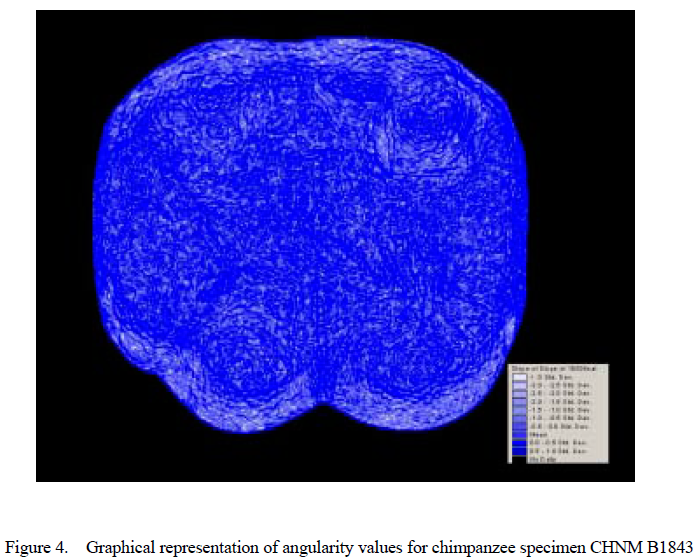
Values for surface slope and angularity were then calculated for each cusp (see Figs. 1– 4, which are published as supporting information on the PNAS web site, www.pnas.org). Slope is defined as the average change in elevation across a surface, whereas angularity is the average change in slope across that surface. Slope refers to the steepness of a cusp, whereas angularity is more a measure of surface jaggedness. The difference between these two variables is analogous to the difference between the sharpness of a knife and its serratedness.
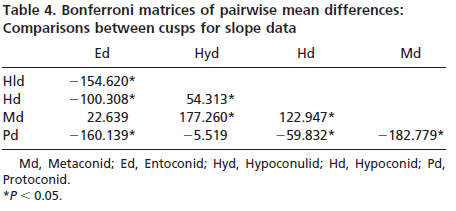
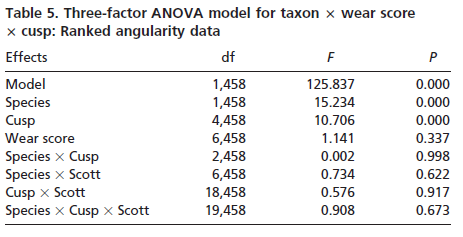
Slope and angularity data were normalized by rank- transformation and analyzed in separate three factor analyses of variance, with taxon, cusp, and wear stage as the factors. This allowed us to determine whether taxa differed in slope and angularity. It also allowed the assessment of whether slope and angularity differed with cusp type or degree of wear. Finally, interactions among the factors would indicate that species differences changed depending on cusp type or degree of wear of the teeth analyzed. Sources of significant variation were determined by using single classification ANOVAs and Bonferroni multiple comparison tests.
Results
Results indicate significant variation in both cusp slope and angularity (see Tables 1– 6). There are no higher-order interactions, and no interactions between any pair of factors in either case. Results for cusp slope indicate significant variation between the two taxa, between cusps and between wear stages. Gorilla specimens have steeper sloped cusps than chimpanzees. Further, lingual cusps have steeper slopes than buccal cusps, with post hoc tests indicating that the entoconid and metaconid have the steepest slopes, the hypoconulid and protoconid have the flattest slopes, and the hypoconid was intermediate. Finally, specimens with higher Scott scores evince f latter cusps than those with lower scores, showing a trend of cusp f lattening with wear. The lack of interactions indicates that degrees of difference in slope between the species and cusp types do not change as the teeth wear.
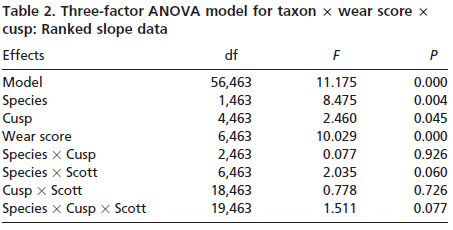
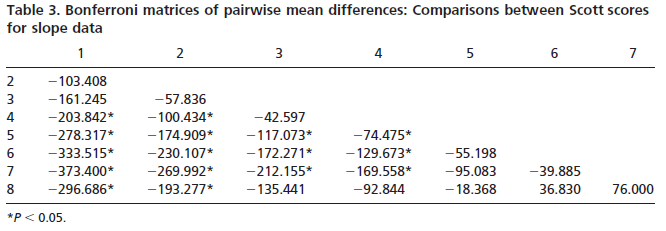
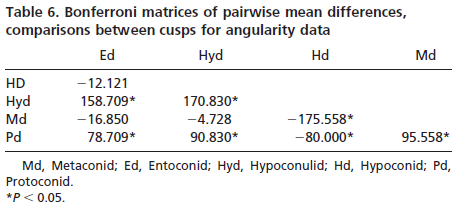
Results for cusp angularity also indicate significant variation between the two taxa and between cusps, but not between wear stages. Gorilla specimens have more angular cusps than chimpanzees. Furthermore, lingual cusps are more angular than buccal cusps, with post hoc tests revealing that the entoconid and metaconid were most angular, the hypoconulid and protoconid are least angular and the hypoconid is again intermediate. The lack of significant differences between wear stages is notable, and suggests that cusp angularity does not change significantly through the wear sequence. The lack of interaction between factors indicates that differences in angularity between the species are comparable for each cusp, and that these differences between species do not change as the teeth wear.
Discussion
This paper outlines a procedure to characterize and compare functional aspects of primate occlusal morphology for worn molar teeth. Traditional dental morphometric studies rely on measurements from cusp tips (4). Although such studies have been effective for separating species by broad diet category, this approach is limited to unworn teeth, as even slight wear obliterates landmarks used for measurement. Dental topographic analysis, on the other hand, models the entire occlusal surface, and is equally effective for studying worn and unworn teeth.
Although we report here on slope and angularity, GIS tools can provide many other useful measurements, such as surface area, relief, topographic aspect, basin volume, drainage patterns(9, 11).
The importance of the ability to include worn teeth in studies of primate dental functional morphology cannot be overstated. First, most fossil species are represented by few if any unworn teeth. This has limited our ability to reconstruct past adaptive radiations of our order using dental functional morphology. Inclusion of worn teeth in such analyses will greatly improve our reconstructions of the diets of past primates. Second, the study of how tooth wear affects form will provide new and important insights into how natural selection acts to maintain efficiency for fracturing foods with specific material properties despite loss of dental tissue with age. This adds a whole new dimension to studies of tooth form and function.
Results presented here for chimpanzee and gorilla teeth offer a glimpse at the potential of this approach. Gorillas are usually classified as folivores, although G. g. gorilla,the subspecies considered here, consumes a considerable amount of fruit when this resource is plentiful (16, 17). Chimpanzees are primarily frugivores, with soft-fruits comprising 70 – 80% of the diets of P.t. troglodytes (12, 13). Where the two subspecies are sympatric at Lopé, the Gabon, gorillas have been reported to consume only 57% of the food species eaten by chimpanzees, suggesting considerable niche separation (13, 18).
As expected given previous shearing crest studies on unworn teeth (19), gorillas have steeper sloped cusps than chimpanzees. Also as expected, cusp slopes decrease as occlusal surfaces wear down for both taxa. The lack of interaction between wear stage and taxon was not, however, anticipated. We expected that as tooth cusps of both species wore down, their slope values would converge. Indeed, the lack of interaction suggests that differences in slope between these species are independent of degree of wear, and are thus maintained throughout the wear sequence. Selection is likely maintaining these differences despite similar degrees of change in slope with wear in each taxon.
Gorillas also have more angular cusps than chimpanzees. Angularity is a measure of jaggedness or change in slope across a surface rather than steepness per se. This finding is consistent with the expectation that a more folivorous form should have more angular surfaces for shearing and slicing tough leaves than a frugivore that needs a f latter, less angular surface for crushing and grinding fruits (20, 21). The lack of interaction between wear stage and taxon suggests that differences in angularity are maintained between species throughout the wear sequence.
It should be noted that cusp slope and angularity relate to different food fracture processes. Cusp slope in primates ref lects angle of approach between opposing teeth during chewing (22).
A steeper slope indicates a steeper approach for slicing or shearing tough foods as opposing occlusal surfaces slide past one another. Lesser relief indicates teeth that come into direct opposition for crushing brittle foods (2). In contrast, angularity relates to variation in shape of a given cutting surface, such as jaggedness or serratedness of a blade. Increasing the angularity or jaggedness of a cutting edge can dramatically change directions of forces acting on a food item, increasing potential damage to that item (23).
The lack of change in angularity with wear for either species is particularly notable, and is among the first demonstrations of the maintenance of a functional aspect of occlusal morphology with wear in primates. This lack of change may be related to slope changes in the occlusal table resulting from angled walls of the pits that form as enamel gives way to softer dentin as the tooth wears. Such a phenomenon would be comparable to that seen in herbivorous ungulates that have complex infoldings and lophs designed to form sharp edges with dentin exposure for shearing and grinding tough foods.
Kay (6) reasoned that thin enamel in primate folivores, such as the gorilla, would lead to sharp edges at the sites of dentin exposure to improve shredding and slicing abilities of the tooth with wear. This is consistent with results presented here, and suggests that cusp angularity might be a critical functional attribute that retains its value despite wear. The difference in angularity values between chimpanzees and gorillas, and the lack of change in this attribute between wear stages further suggests that cusp angularity may be tied to efficiency for food fracture. If so, differences in angularity between taxa, as measured by dental topographic analysis, may be of value for inferring differences in the material properties of the foods to which species are adapted.
We thank Malcolm Williamson, John Dennis, and John Wilson for their help during development of GIS techniques described here. We also thank Mark Teaford for help molding teeth at the Cleveland Museum and Natural History and Bruce Latimer for permission to study specimens in his care. This project was funded by a grant from the LSB Leakey Foundation.